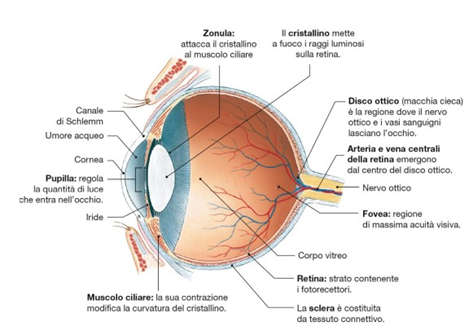
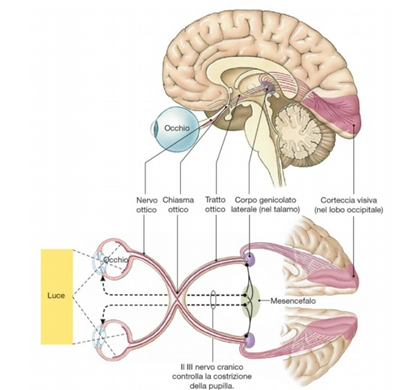
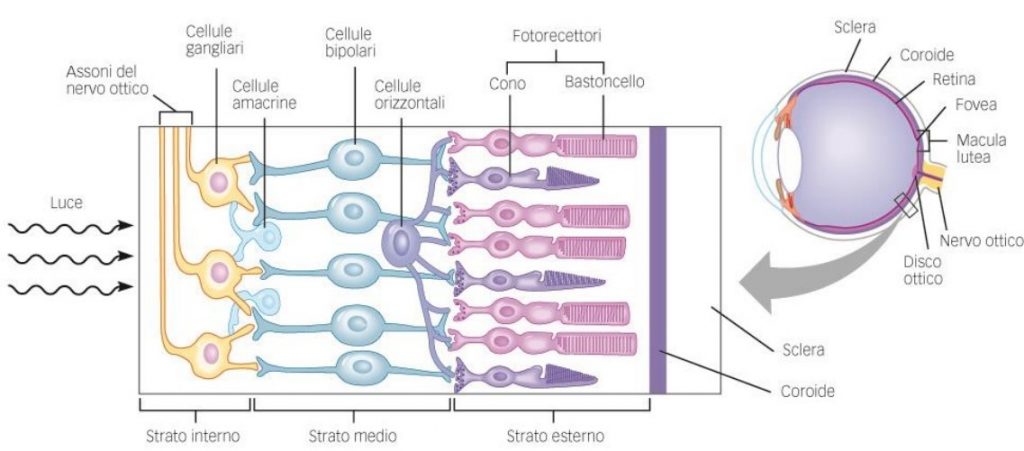
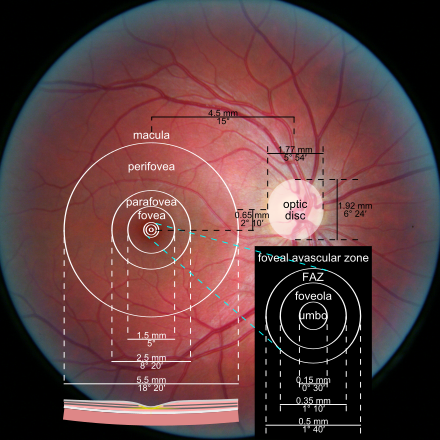
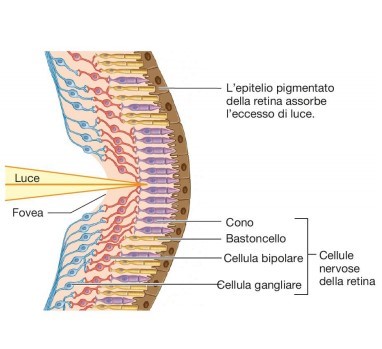
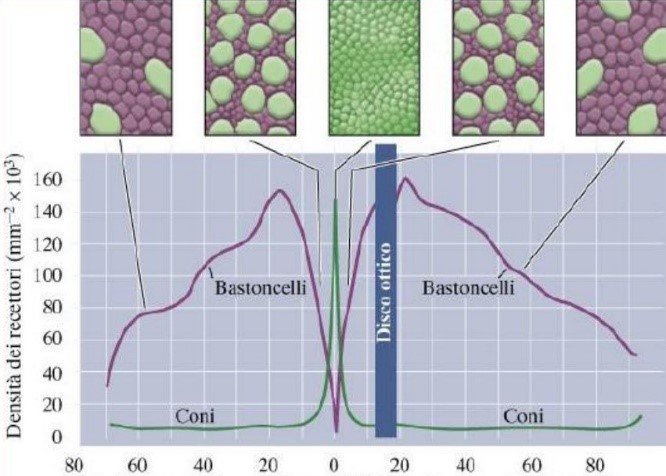
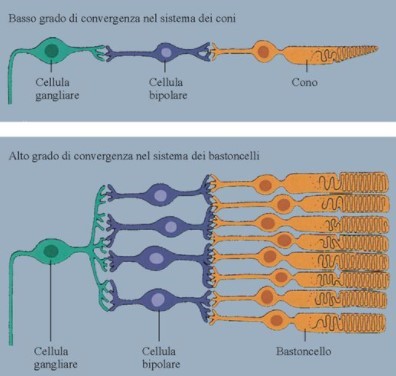
Questo campo recettivo è minimo (può anche ridursi ad un solo cono) nella fovea e nella macula (Fig. 5 in alto); Perciò tali aree retiniche sono caratterizzate da una elevata acuità visiva: meno fotorecettori sono coinvolti, più sarà specifica l’informazione che giunge alla cellula gangliare. La convergenza è invece massima nei bordi esterni della retina, dove una cellula gangliare può avere un campo recettivo formato anche da decine di fotorecettori (Fig. 5 in basso). Questo tipo di organizzazione è coerente alle necessità del sistema nervoso centrale. A livello delle aree corticali visive, infatti, le risorse computazionali sono dedicate agli stimoli su cui è riposta l’attenzione, o che comunque la richiamano, ed essa si sposta inevitabilmente col movimento oculare: gli stimoli significativi tendono nella pratica a trovarsi sempre al centro del campo visivo, dove la risoluzione è massima. Le regioni periferiche del campo visivo, semplicemente, non necessitano di dover garantire uguali prestazioni in termini di risoluzione.
Voi direte, ma tutto questa organizzazione a cosa serve? La risposta è semplice. Noi non dobbiamo solo leggere delle parole e dei numeri, ma dobbiamo muoverci nell’ambiente in condizioni d’illuminazione non sempre uguali e riconoscere le forme e gli oggetti che ci circondano. Per fare questo abbiamo bisogno del riconoscimento del contrasto. La capacità di percepire la minima differenza di luminanza tra due oggetti o aree nello spazio è denominata “sensibilità al contrasto” ed è fondamentale per distinguere gli oggetti dallo sfondo. Il contrasto è necessario per stabilire dove finisce un oggetto e ne inizia un altro, permette il rilevamento di bordi. La zona compresa fra due bordi viene percepita come appartenente ad una superficie uniforme, lo spazio “vuoto” viene riempito. Una differenza di brillanza verrà percepita tra due figure identiche per forma e luminanza in relazione ad una differenza nello sfondo.
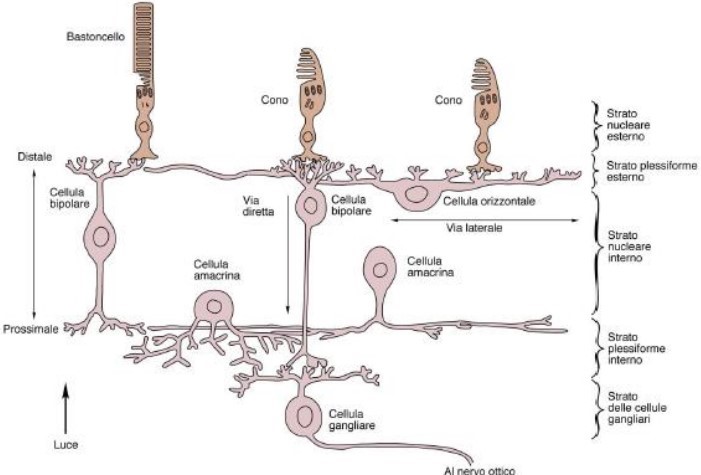
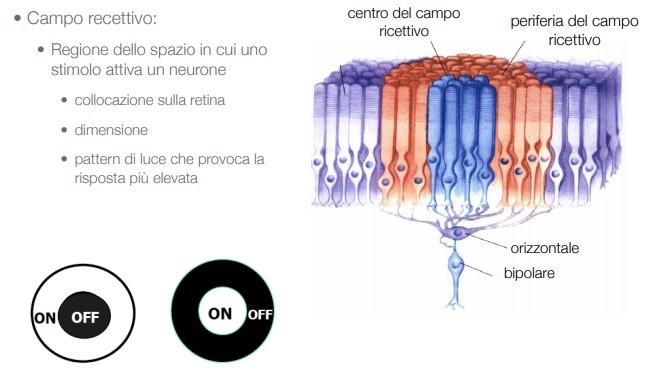
Quindi il processo che rileva lo stimolo luminoso non è opera di un solo recettore (neurone) ma della combinazione di più di questi; l’effetto “corale” della percezione (inibizione laterale) comporta una parziale inibizione del segnale, una distorsione cangiante che appare in movimento. Questo meccanismo nasce all’interno della retina dove le cellule dei fotorecettori e le cellule bipolari, che collegano i fotorecettori alle fibre che raggiungono il cervello, vengono in relazione con le cellule orizzontali, che organizzano e codificano i segnali ricevuti. In pratica, un nucleo di fotorecettori si alimenta in una cellula bipolare, mentre un anello circostante di fotorecettori stimolano le cellule orizzontali a inibire la risposta. (Fig. 6).
Occorre considerare che i campi recettivi retinali hanno una forma pressoché circolare; sono costituiti da una zona centrale e una periferica che circonda la prima. È proprio il contrasto tra l’attivazione (quindi, la luminosità) tra centro e periferia che funge da forte stimolo per le cellule gangliari, innescando una risposta eccitatoria (vale a dire una serie di potenziali d’azione) o inibitoria (senza alcun potenziale d’azione). Un vantaggio significativo di questa soluzione sta nel fatto che gli stimoli deboli possono essere più facilmente percepiti.
Esistono diversi modi con cui le cellule cerebrali addette all’elaborazione dell’immagine si influenzano a vicenda. Uno di questi è proprio l’inibizione laterale: le cellule più attive “spengono” la sensibilità di quelle vicine, rendendole meno attive. Il risultato è che il segnale di un lato del confine tra bianco e nero si amplifica, mentre diminuisce quello che si trova dall’altra parte, creando un contrasto maggiore di quello che esiste in realtà. Ciò permette di vedere le cose in modo più vivido. Tutto questo determina la sensibilità al contrasto che nella realtà è un aiuto che la natura ci ha procurato per distinguere piccole differenze di luminosità (ad es. nello scendere i gradini di una scala in forte penombra.
Riferimenti Bibliografici:
- Snowden, R, Thompson, P, Troscianko, T. Basic Vision: An introduction to visual perception. Oxford University Press, 2006.
- National Eye Institute, Retinal Neurophysiology Section. September 2020.