Senso cromatico
I colori hanno stimolato fin dall’antichità riflessioni sulla loro origine e natura e sul loro ordinamento. I filosofi classici ritenevano che i colori, come i sapori e gli odori fossero qualità degli oggetti determinate dall’apparato sensoriale, ma ad oggi quando si deve dare la definizione di colore occorre prendere in considerazione molteplici fattori. Si tratta di fattori di ordine fisico in relazione alle caratteristiche delle radiazioni luminose e della natura dei corpi, di ordine fisiologico in relazione alle caratteristiche e alle modalità processuali del sistema visivo, e infine di ordine psicologico in relazione all’esperienza del colore. Prima di ogni considerazione sul senso cromatico occorre fare una premessa. E’fondamentale comprendere la differenza fra la luce e i pigmenti colorati presenti nel nostro ambiente terrestre (foglie, alberi, il cielo, il mare, ecc.). La luce con le sue differenti lunghezze d’onda investe i pigmenti che secondo il loro colore ne assorbono una parte e ne riflettono un’altra. E’ quella parte di luce che viene riflessa dai pigmenti che entra nel nostro occhio e attraverso un’accurata analisi neuronale viene tradotta in colore. La visione del colore, quindi, è contemporaneamente sia un’esperienza soggettiva che un’esperienza oggettiva; proprio per questo motivo, per spiegare tale fenomeno, occorre soffermarsi sia sulla luce che sui processi visivi di elaborazione.
Fattori di ordine fisico: la luce
La luce è la condizione necessaria per la visione e soprattutto per la visione a colori. La radiazione elettromagnetica è un’onda che si propaga nello spazio ad altissima velocità e il nostro occhio riesce a percepirne solo una piccola porzione. Ai fini della visione dei colori, l’ampiezza dell’onda influisce sull’intensità luminosa dello stimolo elaborato dal cervello, mentre la lunghezza dell’onda influenza la tonalità del colore percepito.
Nel 1666 Newton dimostrò sperimentalmente che, se si osserva un raggio solare attraverso un prisma di cristallo, esso si scompone in una serie ordinata di raggi colorati aventi differenti lunghezze d’onda (Fig.1). Si vide anche che, collocando due prismi identici in opposizione, si riotteneva il raggio luminoso bianco scomposto precedentemente (Otha and Robertson, 2005).
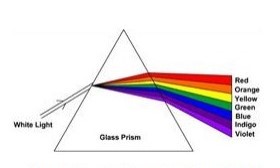
Egli ottenne ciò che noi attualmente chiamiamo spettro visibile. Newton dimostrò che la luce bianca o policroma è costituita dall’insieme delle radiazioni monocromatiche che il nostro occhio percepisce come colore.
Lo spettro elettromagnetico
Lo spettro elettromagnetico è costituito dall’insieme delle radiazioni e comprende l’intera gamma delle lunghezze d’onda esistenti in natura, dalle onde lunghissime alle onde cortissime. Lo spettro è suddiviso convenzionalmente in una successione di bande: onde hertziane, microonde, infrarosso, visibile, ultravioletto, raggi X e γ (P. Mazzoldi, M. Nigro, C. Voci, Elementi di fisica, Padova; EdiSES, 2002). La luce visibile è la banda dello spettro cui l’occhio umano è sensibile. Le differenze individuali possono far variare leggermente l’ampiezza dello spettro che approssimativamente si situa tra i 380 e i 780nm. Alla lunghezza d’onda minore corrisponde la gamma cromatica del blu-violetto, a quella maggiore corrisponde invece la gamma dei rossi (Fig. 2).
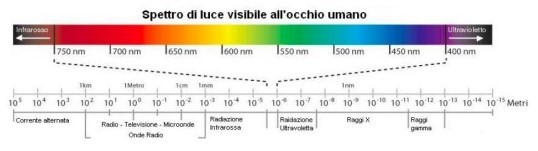
L’ottica è quel settore della fisica che studia i fenomeni connessi all’emissione, alla propagazione e alla rilevazione di un particolare intervallo di radiazioni elettromagnetiche (luce) in grado di produrre stimoli sulla retina dell’occhio umano.
Il campo dell’ottica fisica è spesso allargato alle radiazioni contigue a quelle del cosiddetto spettro visibile e cioè allo studio dell’infrarosso e dell’ultravioletto, in quanto queste radiazioni agiscono sui tessuti che compongono la struttura anatomica dell’occhio. In particolare, la sovraesposizione ai raggi UV causa danni non solo alla pelle ma anche alla cornea, al cristallino e in minor quantità alla retina (B. Allaria, et. all, Protezione dagli UV nelle lenti oftalmiche, Professione Ottico, Medical Evidence, 19, 1-5, 2010).
L’agente fisico che provoca la sensazione visiva è costituito dalle radiazioni elettromagnetiche di lunghezza d’onda comprese tra 380nm e 780nm e questo intervallo, comprende circa 10 milioni di colori percepibili dall’occhio umano (G. Wyszecki, Color, Chicago, World Book Inc., 2006:824 in Discriminate or Diversity di Enrique Ruiz, Maryland, USA, PositivePsycheBiz. Publication, 2009), ma il colore è un’esperienza che dipende da due cose: la luce che gli oggetti riflettono e le proprietà del sistema visivo di chi guarda.
Quando la luce solare illumina un oggetto si possono realizzare i seguenti fenomeni: tutto lo spettro viene riflesso e l’oggetto appare bianco, tutto lo spettro viene assorbito e l’oggetto appare nero, una parte dello spettro viene assorbita e l’altra viene riflessa per cui l’oggetto ci appare del colore della luce riflessa.
Si può ben capire che il colore è funzione delle caratteristiche di assorbimento e di riflessione delle radiazioni luminose da parte dei corpi ma è anche un fenomeno elaborato dai processi che hanno luogo nell’organismo, dalla retina ai centri di codificazione del cervello (Bressan, 2007).
Fattori di ordine fisiologico: dalla retina al cervello
Il complesso meccanismo che porta alla visione e in particolare alla percezione dei colori inizia a livello retinico e termina a livello della corteccia visiva primaria.
La retina è la più interna delle tre tuniche che costituiscono la parete del bulbo oculare. È una membrana sensoriale altamente differenziata negli elementi costituenti, nella quale avvengono, per l’azione delle radiazioni luminose, delle trasformazioni chimiche, fisiche ed elettriche, per mezzo delle quali si origina lo stimolo che, giunto ai centri nervosi, vi determina il fenomeno percettivo della visione. Essendo una struttura neuronale è naturale che sia complessa e i suoi componenti possono esser classificati in molteplici strati come vediamo nella figura 3 (E. Midena., Malattie dell’apparato visivo, CEDAM, 2006).
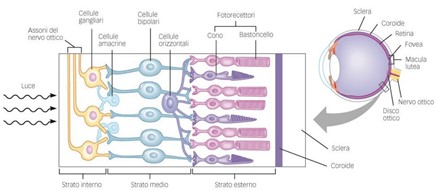
Nella retina sono presenti due tipi differenti di fotorecettori: i coni e i bastoncelli.
Si calcola che nella retina umana siano presenti circa 6 milioni di coni e 120 milioni di bastoncelli. La concentrazione dei primi è massima nella fovea mentre i bastoncelli raggiungono la maggiore densità a circa 20 gradi dalla fovea (Palumbi, 1962; Oyster, 1999). I coni sono deputati alla visione della luce e dei colori e si suddividono in tre tipologie differenti a seconda del fotopigmento localizzato a livello del segmento esterno. Essi sono coni lunghi o rossi, coni medi o verdi e coni corti o blu a seconda della loro sensibilità alle radiazioni luminose (D. Mustafi et al. Structure of Cone Photoreceptors, Progress in Retinal and Eye Research 28:4, 289-302, 2009).
I bastoncelli mediano la visione scotopica e possiedono un solo fotopigmento.
Entrambi i tipi di fotorecettori contengono un pigmento fotosensibile detto porpora visiva o rodopsina, una proteina di membrana costituita da una parte proteica l’opsina e una parte aldeidica, il retinale, che è un cromoforo della vitamina A (L.T. Sharpe, I Used to Be Color Blind, Color Research and Application 26,269-272. 2001).
Nei coni la rodopsina assume alcune varianti atte a conferire ai ricettori la loro particolare specializzazione nella percezione cromatica.
Tali varianti sono:
- L’eritropsina,
- La cloropsina
- La cianopsina
Quando la luce colpisce il fotopigmento, la rodopsina contenuta si divide nelle sue componenti con conseguente sbiancamento del pigmento visivo e liberazione di un neurotrasmettitore chimico. Tale processo dà inizio ai messaggi elettrici dei fotorecettori che alla fine raggiungono il cervello, mentre meccanismi biochimici speciali, rigenerano continuamente le molecole di rodopsina schiarite riportandole alla loro forma originale sensibile alla luce.
Solo quando i segnali della luce e del colore provenienti dall’occhio raggiungono il cervello, vediamo il colore. I coni e i bastoncelli traducono la luce che assorbono in schemi di segnali elettrici che vengono trasmessi attraverso le sinapsi allo strato connettivo di cellule bipolari. Queste mettono insieme le informazioni ricevute dai recettori e le trasmettono verticalmente allo strato successivo, le cellule gangliari, mentre, le cellule orizzontali e i neuroni amacrini distribuiti fra le cellule bipolari trasmettono lateralmente l’informazione (G.K Hung et all. Models of the Visual System, New York, Kluwer Academic Publishers, 109-113,2002).
In particolare, esistono due tipi di cellule gangliari che si differenziano per forma e funzione:
- Cellule Parvo o midget: forniscono informazioni sul colore e sulla forma di un oggetto.
- Cellule Magno o parsol: forniscono informazioni sul movimento e sulle rapide variazioni di luminanza. I loro campi recettivi sono molto ampi ( Zocchi. Principi di fisiologia, Napoli, EdiSES s.r.l., 110-122, 2012).
Il campo recettivo è quella porzione di retina che se stimolata dalla luce produce la massima risposta della stessa cellula gangliare. Esso può essere più o meno esteso e a seconda della sua grandezza fornisce informazioni più o meno precise sulla natura di uno stimolo. In particolare, più sono piccoli i campi recettivi, maggiore sarà la loro densità e migliore sarà la discriminazione di un dato oggetto. I campi recettivi delle cellule M sono molto grandi, mentre quelli delle cellule P sono piccoli ed è questo il motivo della loro alta capacità discriminativa.
Gli assoni delle cellule M e P si riuniscono a formare il nervo ottico, la parte nasale si incrocia a livello del chiasma ottico mentre quella temporale si dirige direttamente al corpo genicolato laterale (CGL) (Fig. 4), struttura suddivisa in sei strati, situata a livello del talamo.
I due strati più ventrali I e II del CGL contengono cellule relativamente grandi e vengono detti strati magnocellulari poiché ricevono la proiezione delle cellule gangliari M. I quattro strati dorsali, detti parvocellulari contengono cellule relativamente piccole e ricevono le proiezioni delle cellule P. Esiste un’ulteriore regione tra gli strati parvocellulari e magnocellulari, detta interlaminare, che contiene neuroni molto piccoli chiamati cellule konio che hanno la funzione di identificare le varie lunghezze d’onda e quindi di riconoscere il colore in maniera grossolana.
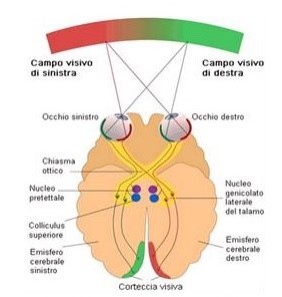
L’informazione visiva, dal CGL prosegue viaggiando tramite le radiazioni ottiche per poi giungere a livello della corteccia cerebrale, e più propriamente nella corteccia visiva primaria (V1) o area 17 di Brodman che si trova collocata per metà nell’emisfero cerebrale destro e per metà in quello sinistro (Gregory, 1990).
Le vie parallele
A livello della corteccia visiva primaria ritroviamo la stessa distinzione presente sia a livello retinico che a livello del CGL per quanto riguarda la suddivisione degli strati in magnocellulari, parvocellulari e koniocellulari.
Di conseguenza si parla di tre vie parallele:
- La via Magno o dorsale codifica il movimento.
- La via Parvo o ventrale codifica la forma e i dettagli degli oggetti.
- La via visiva Konio o blob si ritiene deputata all’analisi del colore.
I campi recettivi dei blob sono ad antagonismo centro-periferia e ad opponenza al colore. Presentano un’alta sensibilità alle lunghezze d’onda, cosa che le altre due vie non mostrano e ciò suggerisce che siano deputate all’analisi del colore dell’oggetto (S.H. Schwartz, Visual Perception, USA, McGraw-Hill, 2004, 3°, 3-162. 2004).
Le cellule della corteccia striata (V1) inviano gli assoni alle vicine aree corticali visive più elevate, che sono le aree V2, V3, V4 e V5. Queste sono coinvolte nell’integrazione delle informazioni visive con gli altri sensi e con la memoria.
L’area V5 è deputata al movimento, l’area V4 al colore e l’area V3 alla forma degli oggetti (C. Casco. Vedere. Torino, Bollati Boringhieri editore, 71-96, 138-159 1992).
Il sistema visivo è quindi un processo assai complesso, caratterizzato da neuroni che rispondono a stimoli sempre più complessi man mano che si prosegue lungo le vie ottiche per ottenere così l’immagine di un oggetto che sia il più rispondente alla realtà per quanto riguarda la forma, le dimensioni e i colori.
Per quanto riguarda la visione dei colori, ad oggi si fa riferimento ad un modello basato su due livelli che concorrono entrambi alla determinazione finale del colore percepito: il primo livello è definito dalla teoria tricromatica, mentre il secondo livello è definito della teoria dell’opponenza cromatica.
Le teorie del colore
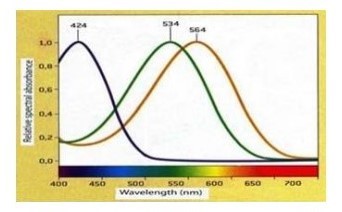
Il meccanismo di percezione dei colori è stato a lungo oggetto di interesse e sono state formulate numerose ipotesi. Le due ipotesi più note sono la teoria tricromatica originariamente proposta da Thomas Young e Hermann von Helmholtz, e la teoria dell’opponenza cromatica proposta da Ewald Hering. La teoria tricromatica fu proposta da Thomas Young nel 1802 e successivamente integrata da Herman von Helmholtz nel 1894. Young supponeva esistessero tre gruppi distinti di fibre nervose, una sensibile al rosso, una al verde e una al blu, basandosi sul fatto che qualsiasi colore poteva essere prodotto da un’opportuna mescolanza dei tre primari. Questa teoria presuppone quindi la presenza di tre tipi di coni, contenenti differenti sostanze fotochimiche che presentano la sensibilità massimale per uno dei colori primari. Non esiste quindi un fotorecettore specifico per ogni singola lunghezza d’onda, ma la visione del colore è data dalla combinazione funzionale di soli tre tipi di coni. Ad esempio, la sensazione del giallo è generata dalla risposta simultanea del fotorecettori rossi e verdi. I tre tipi di cono, denominati L, M e S, si distinguono principalmente per la porzione dello spettro visibile cui ciascuno è estremamente sensibile.
I coni L o rossi sono i più sensibili alle lunghezze d’onda lunghe (assorbimento massimo 564 nm), i coni M o verdi alle lunghezze d’onda medie (assorbimento massimo 533nm) e i coni S o blu alle lunghezze d’onda corte (assorbimento massimo 437 nm) (Fig.5). Queste tre tipologie di coni non sono presenti in egual concentrazione bensì è stimato che gli L siano 2,9 milioni, gli M siano 1,4 e gli S 0,2 milioni per un totale di circa 4,5 milioni di coni per occhio. La stimolazione dovuta a una luce rossa, porta alla visione di questo colore perché vi è una forte eccitazione da parte dei coni che contengono il fotopigmento L, e ad una debole stimolazione degli altri due tipi. Il risultato finale sarà quindi la percezione del colore rosso (C.A. Curcio et al. Distribution and morphology of human cone photoreceptors stained with anti-blue opsin. Journal of. Comparative Neurology, 312:610-624 in The Human Eye, CW. OYSTER, Sunderland, Massachussetts; Sinauer Associates Publisher, 1999).
Ewald Hering propose la teoria dell’opponenza cromatica nel 1878, la quale afferma che la visione cromatica funziona per coppie opponenti di colori: rosso-verde, blu-giallo e nero-bianco.
La risposta data dai coni alle diverse lunghezze d’onda dello stimolo luminoso costituisce l’informazione che poi viene analizzata dalle altre cellule retiniche. La maggior parte delle cellule orizzontali, gangliari e quelle del nucleo genicolato laterale rispondono in maniera differente alla lunghezza d’onda relativa allo spettro visibile: esse sono eccitate da alcune lunghezze d’onda e inibite da altre.
Questi neuroni sono caratterizzati da campi recettivi concentrici a opponenza cromatica semplice (Fig.6) e l’eccitazione del neurone avviene se il centro del campo recettivo viene colpito da un colore, mentre si ha un’inibizione della risposta quando la periferia del campo recettivo risulta colpita dal colore opposto (rosso-verde, giallo-blu). Ad esempio una cellula rosso-on/verde-off (R+/V-) sarà eccitata da luce rossa al centro e inibita da luce verde nella periferia.
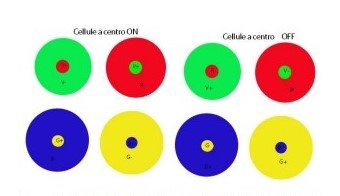
A livello della corteccia visiva primaria troviamo neuroni con risposte più complesse e questi neuroni sono caratterizzati da campi recettivi a opponenza cromatica doppia (Casco, 1992).
La prima considerazione di Hering è che i grigi costituiscono una serie di sfumature che va dal nero più scuro al bianco. Tutti gli altri colori si possono ordinare in altre quattro serie di sfumature: dal rosso al blu, dal blu al verde, dal verde al giallo e dal giallo al rosso. Gli estremi di queste quattro serie sono i cosiddetti colori fondamentali.
Poiché i colori considerati sono quattro, cioè rosso, verde, giallo e blu la teoria è anche nota come teoria dei quattro colori o tetracromatica.
Ai tempi di Hering questa teoria non era compatibile con la teoria tricromatica, e la controversia tra Helmholtz e Hering durò diversi anni.
Da alcuni studi sui pesci (T. Tomita et al. Spectral response curves of single cones in the carp, Vision Research. 7,519-531.1967), sulle scimmie (R.L. De Valois. Analysis of response patterns of LGN cells”, Journal of the Optical Society of America 56:7, 966- 977.1966) e sugli uomini (P.K. Brown et al. Visual pigments in single rods and cones of the human retina, Science 144, 45-51. 1964), è emerso che le due teorie sono entrambe corrette ma si riferiscono a due momenti diversi del processo visivo.
L’uso di tecniche avanzate e i progressi nel campo dell’elettrofisiologia, hanno permesso di misurare l’assorbimento spettrale dei singoli coni presenti sulla retina umana e su quella della carpa. I risultati ottenuti mostrano chiaramente la presenza di tre tipi di coni con picchi di assorbimento relativi alle lunghezze d’onda di circa 450±3, 525±3, 560±3 nm. Le ricerche sulle scimmie, effettuate dal neurofisiologo statunitense R.L. De Valois, hanno dimostrato a livello delle cellule orizzontali, bipolari, gangliari e del corpo genicolato laterale, la presenza di campi recettivi a opponenza cromatica centro-periferia. Questi rispondono in maniera differente alle variazioni di luminanza e di lunghezze d’onda.
La risposta tricromatica è quindi presente nei coni mentre la risposta cromatica opponente riguarda gli stadi successivi dell’elaborazione dello stimolo luminoso e cioè le cellule orizzontali, gangliari, il CGL e la corteccia visiva (L. Maffei e L. Mecacci. La visione, dalla neurofisiologia alla psicologia, Milano, Mondadori, 97-116.1979).
Fattori di ordine psicologico: il senso cromatico e i pigmenti.
Il senso cromatico è la capacità di distinguere correttamente i colori del mondo che ci circonda e che ci permette di riconoscere le caratteristiche proprie dell’oggetto valutandone il volume, la pesantezza e il probabile materiale di cui è composto. Tre distinte qualità fisiche e psicofisiche ci permettono di distinguere un colore da un altro. Esse sono: tono, saturazione e intensità (E. Miotto, Luce e colori, Milano, Fenice 2000, 1994).
- Il tono o colore propriamente detto, caratterizzato dalla lunghezza d’onda dello spettro.
- La saturazione, indica la purezza o l’intensità di un colore. Un colore saturo quindi non contiene parti di nero o bianco.
- L’intensità o luminosità è il carattere che permette di distinguere un colore luminoso da uno opaco.
La materia è composta da una struttura molecolare che reagisce alla luce in modi diversi, infatti, secondo la propria composizione, assorbe alcune radiazioni e ne respinge altre. Ogni sostanza o superficie che ci appare colorata, sottrare alcune radiazioni alla luce e trasmette per riflessione tutte le altre, in base alla propria struttura molecolare. Ciò che viene percepito dall’occhio umano come colore, sono le radiazioni riflesse, perché i corpi materiali non sono né luminosi né colorati ma sono percepiti come tali solamente dall’occhio umano. Un corpo nero assorbe tutte le radiazioni elettromagnetiche visibili, mentre un corpo bianco le respinge tutte. Un corpo giallo, per esempio, assorbe le radiazioni comprese nella zona spettrale del viola e riflette nella zona degli arancioni e dei verdi. Un occhio normale è in grado di percepire non solo i colori fondamentali puri ma anche le varie combinazioni cromatiche che derivano dalla mescolanza di essi eseguita in varie proporzioni.
La luce bianca o policroma è costituita da differenti radiazioni monocromatiche che compongono lo spettro del visibile e che il nostro occhio percepisce come colore. L’uomo però non è in grado di distinguere le componenti della luce, ma ne percepisce la sintesi.
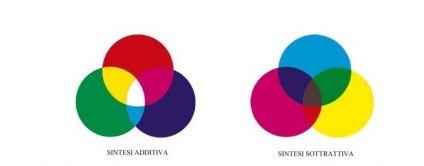
Nella sintesi additiva (Fig.7) i colori sono suddivisi in:
- Colori primari: rosso, verde, blu
- Colori secondari: giallo, ciano, magenta.
Nella sintesi sottrattiva (Fig.7) i colori sono suddivisi in:
- Colori primari: giallo, ciano, magenta.
- Colori secondari: rosso, verde, blu.
Nella sintesi additiva la somma di più onde visibili, che ai nostri occhi appaiono colorate, dà il bianco; mentre nella sintesi sottrattiva la somma di più pigmenti colorati dà il nero (L. De Grandis, Teoria e uso del colore, Milano, Mondadori, 2000).
Grazie alla conoscenza della fisiologia della visione dei colori, ci si è potuti dedicare allo studio delle anomalie della percezione cromatica e allo sviluppo di test diagnostici utili per la gestione e la compensazione di questi difetti mediante l’utilizzo di filtri specializzati.
Occhiocapolavoro
Dott. Giuseppe Trabucchi – Medico Chirurgo – Specialista in Clinica e Chirurgia Oftalmica
P.IVA 02128970031 – C.F. TRBGPP59D30E463K
Iscrizione Ordine dei Medici Chirurghi di Milano n. 25154